
Clinical Oncology Research Journal
Research Article
Verification of MRI MAGAT Polymer Gel Dosimetry in Clinical Radiotherapy Applications
Safaeian GH1*, BahreyniToossi MT2, BahreyniToossi MH2, Feyzi A3, Bayani SH2, Heravian Shandiz M1
1Department of Radiation Technology, School of Paramedical Sciences, Mashhad University of Medical Sciences, Mashhad, Iran
2Medical Physics Research Center, Faculty of Medicine, Mashhad University of Medical Sciences, Mash- had, Iran
3Department of radiology, School of Medicine, Mashhad University of Medical Sciences, Mashhad, Iran
(ORCID:
GhorbanSafaeian: 0000-0001-9409-3110
Mahdi HeravianShandiz: 0000-0002-5199-3503 )
*Corresponding author: Safaeian GH, Department of Radiation Technology, School of Paramedical Sciences, Mashhad University of Medical Sciences, Iran, E-mails: SafaeianGh@mums.ac.ir
Citation: Safaeian GH, BahreyniToossi MT, Feyzi A, Bayani SH, Nouri M (2019) Verification of MRI MAGAT Polymer Gel Dosimetry in Clinical Radiotherapy Applications. Clin Oncol Res J: CORJ: 100001
Received Date: 13 March, 2019; Accepted Date: 20 March, 2019; Published Date: 27 March, 2019
Abstract
Background: The purpose of this work was to study the ability of MRI normoxic polymer gel dosimetry system.
Methods: To accomplish of this study, 2 liters of the normoxic MAGAT polymer gel was composed. A Perspex phantom and five calibration test tubes were also prepared. The test tubes were filled in with the gel. This phantom was then CT scanner and dose plan was product. The gel phantom and the test tubes were then irradiated. Prior to irradiation, MR scans were performed to measure the background value of R2 of the gel. Immediately after irradiation new images of the gel phantom (and calibration test tubes) were obtained using the MR scanner. Finally, from the MRI images in MATLAB environment R2 maps were calculated.
Results: In this study, and in point center (PC) the difference between the treatment planning system TPS and gel dosimeter data was 1.15% (SD = 1.8%). Dose sensitivity and dose resolution of MAGAT gel dosimeter were 5.033 S-1Gy-1 (R2 =0.9953), 1.974 Gy respectively.
Conclusion: In this work, the TPS calculations compared with polymer gel dosimeter measurements and found the dose distributions calculated with the TPS is in very good agreement with the Polymer gels measuring.
Keywords: Calibration; Gel dosimetry; Magat; Normoxic; Radiotherapy
Introduction
In radiotherapy there is a great need for accurate determination of the adsorbed dose of tumor tissue as well as to healthy organs at risk. The absorbed dose delivered to the planning target volume (PTV) should be %5 of the stated dose (Other authors suggest %3) [1]. To accomplish this goal, most treatments are executed according to a calculated plan. In clinical cases verification of the calculated dose plan by measurements is often complicated. This is especially true if the treatment comprises server al beams of different field and radiation qualities and if the beams impinge on an irregularly shaped body section containing various kinds of tissues or cavities. One method to verify clinical treatments is to carry out measurements using thermoluminescence (TL) dosimeters in patient-like phantoms [2]. This method is limited with respect to the absorbed dose in a limited number of points.
Furthermore, the detectors may disturb the radiation beam or their signal be dependent on the radiation qualities used as well as the direction of the incident radiation. These are problems which TL dosimeters share with most other dosimetry systems such as diodes and ionization chambers. Most conventional dosimetry techniques that mentioned above are incapable of 3D measurements. Polymer gel dosimeters are able to measure dose distributions for several beams and different beam qualities (e.g., photons and electrons) [3]. The purpose of the present study was to study the ability of MRI normoxic MAGAT polymer gel dosimetry system as a tool to verify the calculated dose distributions in clinical radiotherapy (Prostate cancer).
Material and Methods
For verification of absorbed dose distribution using the MAGAT gel dosimeter initially, a cylindrical Perspex phantom simulating a patient and a number of calibration test tubes were designed and composed (Figure 1). 2 liters of the normoxic MAGAT polymer gel under normal atmospheric conditions was prepared according to the reference method [4]. Five calibration test tubes and the Perspex phantom were filled in with the prepared gel. The phantom was then CT scanner and a dose plan was generated using the obtained CT data (Figure 2). Prior to the irradiation, MRI scans of the phantom was taken to measure the background value of R2 (Figure 3 and Table 1, 2). The gel phantom was irradiated according to four field (Box) technique protocol. The calibration test tubes were also irradiated by a cobalt-60 tele therapy unit (Figure 4). Immediately afterthe irradiation, new images of the phantom and the calibration test tubes were acquired using the MRI scanner. Finally, using the MRI images in MATLAB environment R2 maps were calculated.
The Phantom, Test Tube and Gel Preparation
In this study, a special cylindric Perspex phantom was designed to investigate dose distribution in external beam radiotherapy of urinary bladder. Perspex was chosen because it is relatively soft tissue equivalent and cheap, easily machined and useful for constructing phantoms of varying shapes. Thickness the walls of phantom was 1cm Perspex and the dimensions were 15 cm in diameter, 15 cm in height and 1200 ml capacity. This phantom is composed of 3 separate units. The central one contains gel dosimeter (1200 ml) and the other two contain the pure water (19 x 25 cm, and 3600ml capacity). For calibration purpose, relatively small Pyrex tubes were designed with 2mm wall thickness, 1.5mm diameter, 8cm height and 15.5 ml capacity (Figure 1).
"MAGIC" which stands for "Meth acrylic and Ascorbic acid in Gelatin Initiated by Copper", was the first normoxic polymer gel proposed [4]." MAGAT" polymer gel is one of the most sensitive radiation of the normoxic gels, that had been used in this study. The fabricating of the new gel followed the same manner as reported for MAGIC. For fabricating of 2 liters of this gel we used; 160 gr gelatin (type A, 300 bloom) 662 gr THPC agent, 100 gr meth acrylic acid (MAA) and 1740 ml HPLC water.
Calibration and Irradiation
In this study a standard calibration multi-tube with five test tubes were employed. A tele-therapy Cobalt-60 machine (Theratron 780- AECL) was used to irradiate the test tubes laterally when they were horizontally fixed inside a water tank at the depth of 5 cm with a source-to-surface distance (SSD) of 80 cm (Figure 4). One tube was left Unirradiated while the others were irradiated to doses of 2,4,6,8 Gy. Front and back surfaces of the tubes were marked by adhesive stickers and the test tubes were irradiated with their front side facing up to the beam. This marking was later used for positioning the imaging slice at the middle of the test tubes between the front and back surfaces where the gel was exactly at the depth of 5 cm during irradiation. The gel phantom was irradiated according to four field (Box) technique protocol that come followed in Table 1.
CT and MRI Imaging, Data Post-Processing
CT scanning of the phantom was performed by a Siemens Somatom Plus-s, 64 slice, and rotate-rotate machine of the third generation. MRI scans of the phan- tom and the calibration test tubes were also taken to extract the spin- spin relaxation rates and R2 maps (Table 2). Images were obtained in a plane in the middle and across the longitudinal cross section of the Phantom and test tubes. 16 MRI images for the phantom and 32 images for the test tubes were obtained. MRI images were transferred in DICOM format to a personal computer for further image processing. Image averaging and background subtraction were performed using the special software (is named R2 calc), that is performed in MATLAB™ environment. Also, an Adaptive and a Median filter with different pixel size masks were also applied on the final polymer gel dosimeters images.
Results
5.1 Calibration Results
The calibration results are followed in Table 3, 4 and Figure 6. Based on these results the dose sensitivity of MAGAT gel dosimeter was 5.033 S-1Gy-1 (R Square, (R2),=0.9953). Dose linearity and dose resolution of this normoxic gel were determined also, and were 0 -8 Gy, 1.974 Gy respectively.
Radiotherapy Procedures Results
Four field technique normally applied for treatment of cancer of urinary bladder was implemented to a gel dosimetry phantom. Gel dosimetry and treatment planning system (ALFARD, Version 4.46.7 SPL WP) were employed to obtain the absorbed dose distribution. Homogenous dose distributions were produced by both techniques. R2 maps and dose contours of the MAGAT gel phantom were obtained in different views after using a adaptive and median filter (5x5, 10x10 and 2x10 masks) (Figure 7, 8, 9, 10, 12). R2 profile of MAGAT gel phantom in different views also determined (Figure 11,13). Dose Volume Histograms (DVHs) and the normalized isodose contours for gel and TPS (ALFARD) system obtained and compromised (Figure 16).
ICRU (42) guidelines researches were implemented to comparing and evaluating these dose distributions [6, 7]. The ICRU (42) hasrecommended that the computed dose should deviate from measured dose by less than 2% [6]. In this study and in the region of interest (at the central slice of the phantom, point center) the difference in the dose obtained by gel dosimetry and TPS is 1.15% (SD = 1.8%), and in penumbra region was 2- 4%.
Discussion
In this study dose distribution of a clinical treatment procedure produced by MRI normoxic polymer gel dosimetry and TPS were investigated. This was accomplished by pixel-by-pixel, isodose and dose volume histogram (DVH) comparison. Based on results, dose-integrating capacity of the gel dosimeter was demonstrated. A good agreement was also found between the data obtained by the two methods employed. The TPS calculated data were in very good agreement with the distribution measured by polymer gel dosimeter. How ere, in a beam abutment region (for the penumbra of the lateral scatter contribution), larger dose difference was found (DD =2- 4 %). The new polymer gel that was fabricated in this study, was also found to have a higher dose sensitivity compared to other normoxic gels.
In order to make a comparison with other studies, we have used one qualitative and one quantitative indicators, respectively Dose Deviation (DD) and Distance To Agreement (DTA)
Conclusion
Polymer gel dosimetry has been developed into a totally non-invasive and non- destructive dosimetry method, since the dosimeter gel phantom itself forms the detector. The gel dosimeter is capable of measuring dose distributions from several beams and beams of different radiation qualities (e.g. photons and electrons) in all parts of the dosimeter volume [7]. These properties imply that the gel dosimeter may be used to verify a dose plan. The purpose of this work was to study whether computerized planned clinical treatments could be verified using the gel dosimeter and to examine possible explanations to deviations found between the calculated and measured dose distribution. Based on the results of this study, the gel dosimetry method was proven to be a useful tool for radiation treatment planning verification.
Acknowledgments
The authors would like to thank research council president for research affairs and office of vice of Mashhad University of medical sciences for their approval and financial support of this work. Thanks, are also given tomembers of staff and technicians of radiation therapy department of Ghaem hospital; CT-scan and MRI departments of Razavi hospital of Mashhad.

Figure 1: Phantom and calibration test tubes to be used (in this study).
Figure 2: CT scanner system and DICOM image of phantom.
Figure 3: MRI scanner system used in this study.

Figure 4: Calibration set up under the tele Cobalt unit in multi tube method therapy.
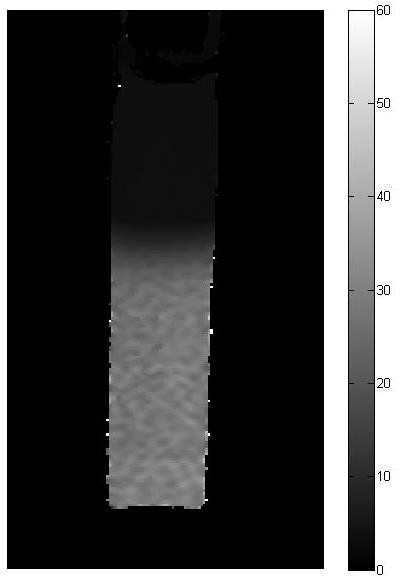
Figure 5: MRI slices of calibration test tubes.
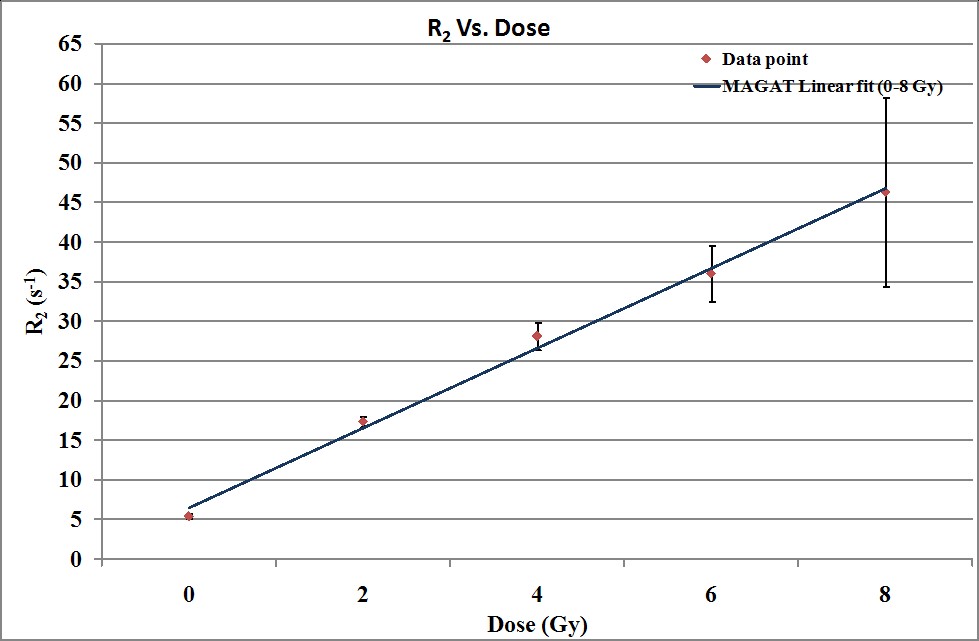
Figure 6: Dose response curve of MAGAT gel dosimeter was built in this study.

X(PiXel)
Figure 7: The coronal view of the pre-irradiated gel phantom.
X (Pixel)
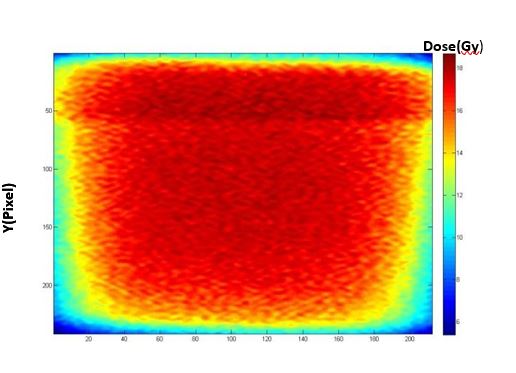
Figure 8: The coronal view of the post-irradiated MAGAT gel phantom (Box technique).
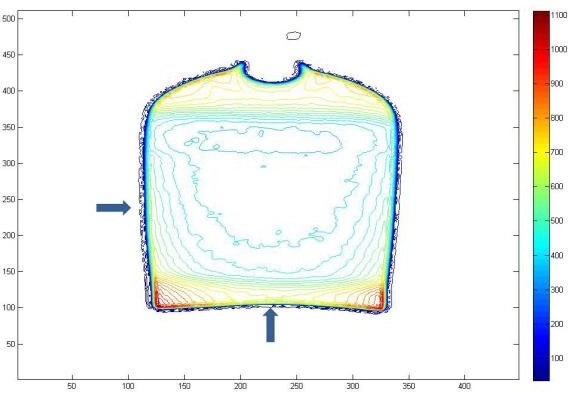
Figure 9: R2 contour of MAGAT gel phantom in coronal view after Using an adaptive filter (10 x 10 mask) (Box technique).
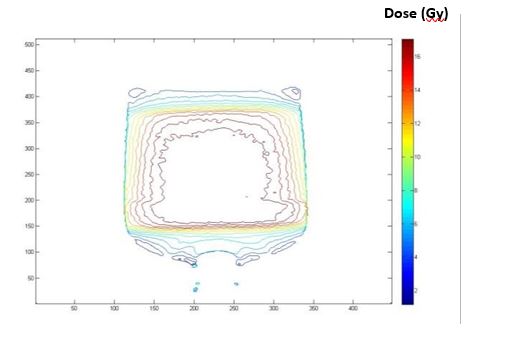
Figure 10: Dose contour of MAGAT gel phantom in coronal view after Using a adaptive filter (10 x 10 mask) (Box technique).
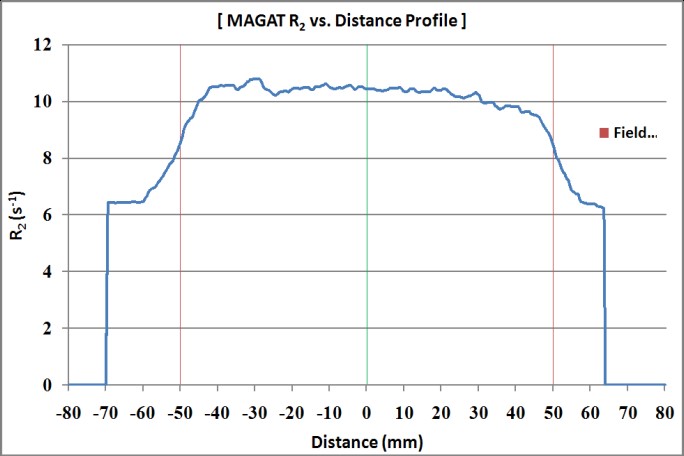
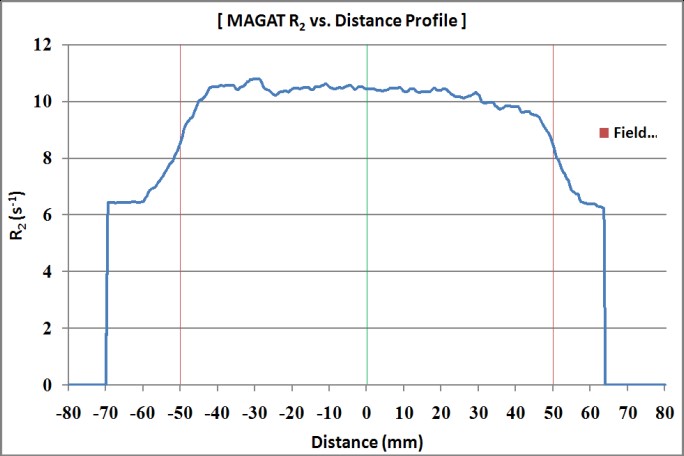
Figure 11: R2 profile of MAGT gel phantom in coronal view (Boxtechnique).
Z (Pixel)
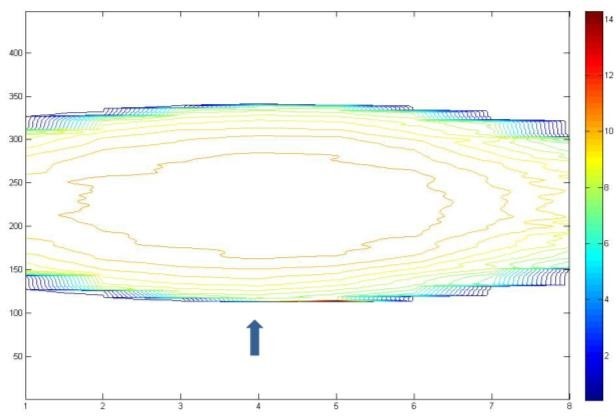
Figure 12: R2 contour of MAGAT gel phantom in Trans axial view after Using a adaptive filter (2*10 mask) (Box technique).
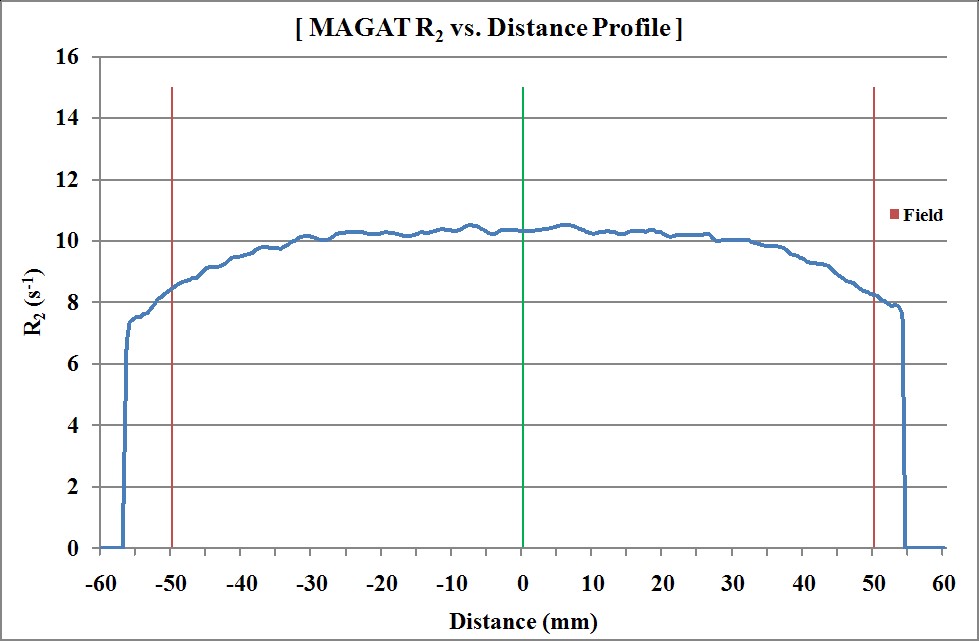
Figure 13: R2 profile of MAGAT gel phantom in Trans axial view (Box technique).
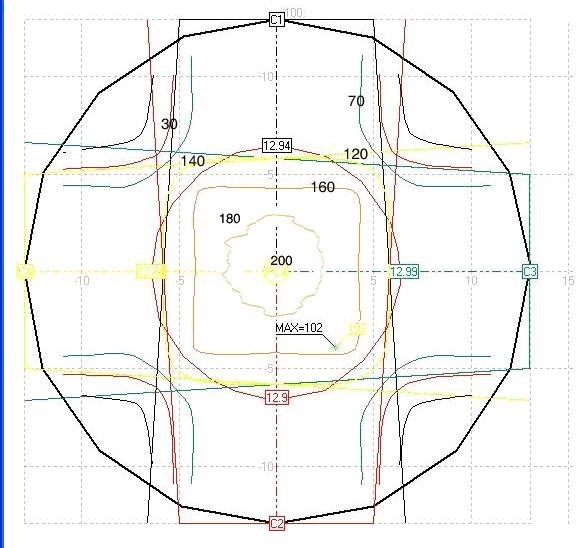
Figure 14: Isodose curves by ALFARD treatment planning system (Box technique).
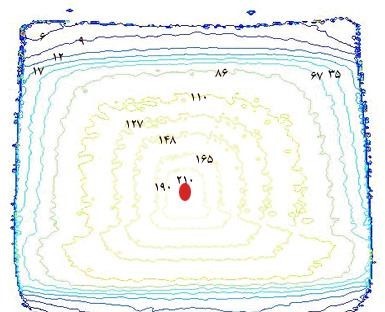
Figure 15: The normalized isodose contours by MAGAT gel Dosimetry system in this study (Box technique).
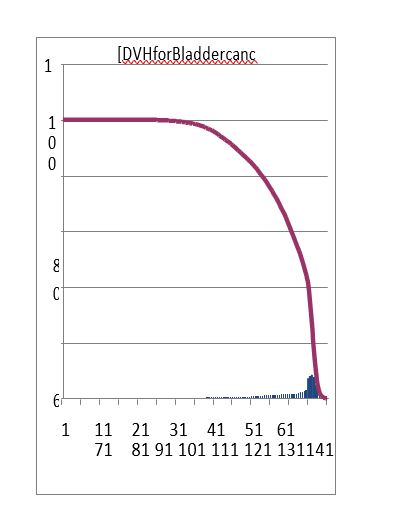
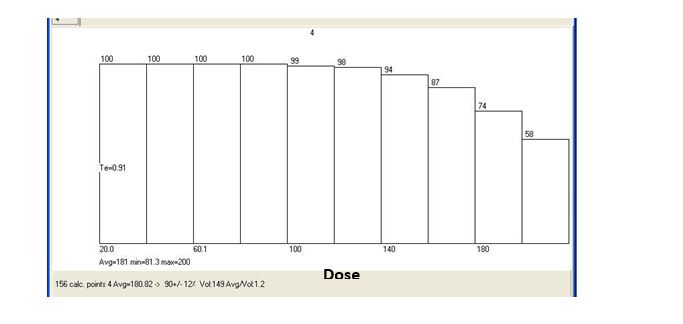
Figure 16: Dose Volume Histogram (DVH) of MAGAT gel phantom and TPS in Box technique (a: gel dosimeter and b: ALFARD TPS).
|
Treatment |
(SSD) |
Field size |
Prescribed dose fromone |
Technique |
|
time (Min) |
(cm) |
(cm) |
field (cGy) |
|
|
1.8 |
80 |
10 x 10 |
50 |
Four field (Box) |
Table 1: Irradiation parameters for prostate cancer treatment (Box technique).
|
Scanner parameters |
Scanner data |
|
Scanner type |
Siemens Avanto (Germany) |
|
Field strength |
1.5 Tesla |
|
RF frequency |
65 MHz |
|
Coil used |
Quadrature head coil |
|
Pulse sequence |
Multi-spin echo (CPMG) |
|
TR (ms) |
3000 |
|
TE (ms) |
22-704, TEn= 32(TEi =22) |
|
FOV Read (mm) |
260, 230 |
|
Matrix size |
512?512 |
|
Slice thickness (mm) |
4 |
Table 2: MRI Scanner Parameters employed in this study.
|
Number of test tubes |
Dose (Gy) at the central axis of the test tube |
Irradia- tion time (min) |
R2 (at the central axis of the test tubes and sagital plan) (1/s) |
SD (?cal) |
|
Blank |
0 |
0 |
5.35 |
0.34 |
|
2 |
2 |
3.05 |
17.36 |
0.64 |
|
3 |
4 |
6.11 |
28.12 |
1.66 |
|
4 |
6 |
9.16 |
36.03 |
3.53 |
|
5 |
8 |
12.22 |
46.35 |
11.90 |
Table 3: Results of multi-tube calibration (in this study).
|
Comments |
Special indexes |
Dose-response equilibrium |
Features / Linearity(Gy) |
|
Coefficients (with 95% confidence bounds |
SSE*=4.7886 R Square=0.9953 |
Linear fitted model Polynomial: R2(D) = P1*D + P2 P1 =5.033 (4.397, 5.669) P2 =6.509 (3.395, 9.624) |
0-8 |
Table 4: Property of MAGAT gel in multi-tube calibration (in this study).
Citation: Safaeian GH, BahreyniToossi MT, Feyzi A, Bayani SH, Nouri M (2019) Verification of MRI MAGAT Polymer Gel Dosimetry in Clinical Radiotherapy Applications. Clin Oncol Res J: CORJ: 100001.
