
Open Access Journal of Agriculture Research
(ISSN 2651-9003)
Research Article
Assessment of Environment and Genetic Impacts On the Variability of Morphological Traits of Apricot Germplasm from Various Genetic and Environmental Resources in Morocco
Ayour J1,2*, Alahyane A1, Elateri I3, Alfeddy MN4 and Benichou M1
1Food Sciences Laboratory, Faculty of Sciences Semlalia, Cadi Ayyad University, M-40090 Marrakech, Morocco
2University of Avignon and the Pays du Vaucluse, UMR408 Safety and Quality of Vegetable Products, F-84000 Avignon, France
3Laboratory of Molecular Biology and Food, Center of Analysis and Characterisation, Faculty of Sciences Semlalia, Cadi Ayyad University, P.O. Box: 2390, 40090 Marrakech, Morocco
4Laboratory of Phyto-Bacteriology, Plant Protection Research Unit - Regional Center for Agronomic Research, INRA, BP 533, Marrakech 40000, Morocco
*Corresponding author: Jamal Ayour, Food Sciences Laboratory, Faculty of Sciences Semlalia, Cadi Ayyad University, M-40090 Marrakech, Morocco, Tel: +33 6 99 75 47 64; Email: jamal.ayour@educagri.fr
Citation:Ayour J, Alahyane A, Elateri I, Alfeddy MN, Benichou M (2019) Assessment of Environment and Genetic Impacts On the Variability of Morphological Traits of Apricot Germplasm from Various Genetic and Environmental Resources in Morocco. Open Acc J Agri Res: OAJAR-100025
Received date: 21 December 2019; Accepted date: 24 December 2019; Published date: 31 December 2019
Abstract
Agro-morphological traits were evaluated for 92 Moroccan apricot accessions issued from different geographical sites, using 34 morphological qualitative and quantitative characters. Strong correlations between studied traits were observed, especially between fruit and stone weights, fruit dimensions, flesh firmness and color traits. Significant variations (p ≤ 0.001) were observed among studied parameters which can help differentiate between different apricot accesions. The analysis of structure was able to show that leaf and fruit dimensions as well as the skin color of fruit represent the discriminating parameters of studied apricots. No clear morphological structure was displayed according to geographical origin compared to apricot genotype. However, two apricot categories have been identified: an apricots group grown under arid conditions with a relatively long winter period and lower temperatures. And a second group grown under Saharan climatic conditions with a short winter period and high temperatures starting in early spring. This is the first report on the morphological analysis of the genetic diversity of Moroccan apricot accessions. This study would provide a solid and genuine basis for effective management and sustainable use of apricot genetic material in future breeding programs in the Mediterranean region.
Keywords:Accession; Environment; Geographic Origin; Morphological Traits; Variability
Introduction
Prunusarmeniaca L. is one of the most important Prunus species grown in the world. More than 80% of the world’s apricot production is restricted to the Mediterranean region, mainly in Turkey, Italy, Greece, France, Morocco and Spain. The apricot production of these countries is based mainly on many local cultivars (FAO, 2013). The apricot is known by its great variability, described by its ecological requirements, its histological features and agronomic conditions as well. Therefore, many differences are found in the quality of the apricot fruits. This shows the importance role of selecting and classification of apricot’s genotypes based on high-quality fruit attributes which would satisfy the consumers [1]. Moreover, the apricot quality is affected by agronomic and ecological factors [2] as well as pomological traits [3]. These characteristics depend on the biological properties of the tree [4] and especially on the fruit and leaf morphological attributes [5].
Morphological characterization is useful for cultivar identification, selection, differentiation and germplasm management. Morphological traits are essential in the establishment of programs designed for the selection of genotypes and they are qualitative traits for markets requirements [6]. Several studies have analysed the variability of apricots, and interesting cultivars have been identified and used to characterize the genetic diversity of apricot on one hand [7] and to generate interesting apricot selections through breeding programs on the other hand [8]. The apricots are distinguishable based on fruit characteristics, such as size, weight, skin colour and shape being the most discriminatory parameters. Indeed, the characterization of accessions is vital to avoid the loss of diversity and to identify and preserve potential valuable traits and characters that contribute the most to the total diversity. Additionally, the agronomic and phenological traits are important for the improvement of the plant’s response to agricultural management and crop breeding particularly in uncertain future climate scenarios [9].
Currently, the estimated area of cultivated apricot trees in Morocco is of the order of 12000 ha with an average production of about 1.2 million tonnes and an average yield of 11 tonnes per hectare. The Marrakesh region is one of the most important cultivated apricot growing areas in the country. The traditional varieties are almost the only source of production. Various renovations have occurred in recent years to introduce new interesting cultivars to meet market requirements and satisfy consumers and industry demands. The National Institute for Agricultural Research in Marrakesh (INRA) has undertaken a long research program targeting areas of traditional cultivation of several fruit species including the apricot fruit. This work of prospecting, collecting and evaluating of the local plant material, made possible the selection of several genotypes to provide them to the professionals by dint of their performances and advantages. They also helped to accumulate important data of pomological description and establish a collection of a wide range of new apricot genotypes in the experimental fields of the INRA. Significant efforts have been made previously to test the performance of Moroccan apricot cultivars [10]. However, to our knowledge, no research has comprehensively addressed the relationships among morphological and phenotypic traits of apricots fruit and leaves by multivariate statistical methods which involve the simultaneous analysis of several variables and provide more useful and accurate information.
The aim of this study is to evaluate morphological and biometric traits of fruits and leaves among ninety-two Moroccan apricot accessions and to evaluate the impacts of the environment and the gographic area on the genetic diversity of apricots. The studied cultivars have been analyzed according to their agronomic performances and geographical origins. This study aims also to investigate morphological variability and its implication in the genetic diversity of Moroccan apricot. We have used principal component analysis to study correlations among studied traits and to establish relationships among cultivars as a tool for germplasm description and selection. This will be the first study of the apricot’s morphologic diversity issued from different Moroccan regions by using a methodology allowing us to compare our results with those of other groups in the Mediterranean basin.
Material and methods
Field survey
A total of 92 apricots accessions were analysed in this study. The accessions were maintained in experimental orchard of the National Institute of Agricultural Research (INRA, Regional Center in Marrakech 30°21’08.4” N, 9°30’29.0” W). They were collected from different origins in Morocco, could be divided in different populations according their geographical sites and geographical regions (Table 1).
Among these accessions, 50 were collected from 9 oasis agroecosystems (Figure 1) located in the four main geographical regions in the south-east of Morocco, each with distinct ecological conditions (Figure 1; Table 1): Ziz Valley [Er-Rich (12), Aoufous (5) and Goulmima (4)]; Draa Valley [Skoura (7) and Agdez (5)]; Dades Valley [Boumalen (8) and El-KalaaM’Gouna (4)] and Moulouya Valley [Guercif (1), Outat El-Haj (4)]. In addition, 9 cultivars grown at the Ain Taoujdate Experimental Station (INRA-Meknes, Morocco) were studied, including eight accessions identified by Barbeau and Bouami (1980) (Agmat 5, Khorb 6, Man 15, Marouch 3, Marouch 4 and Marouch 16) and three introduced cultivars (Delpatriarca, Gelitano and Maoui). Furthermore, 33 additional cultivars grown on the site of Marrakech were analyzed.
Morphological data: Morphological evaluation was undertaken during both 2014 and 2015 seasons. Representative samples were collected as follows:
The morphological characterization of the 92 cultivars was carried out using the UPOV descriptor (2012). Leaf characterization was carried out in March and April, whereas fruit morphological characterization was assessed during May and June for selected apricots at the commercial maturity stage. Twenty-three qualitative morphometric variables and eleven quantitative morphometric parameters were measured. Overall, 34 quantitative and qualitative morphological traits were assessed (Table 2).
Statistical analysis: Statistical analysis was conducted using SPSS analysis software version 19.0. A data matrix of quantitative and qualitative variables was used for statistical calculations. Descriptive statistics represented the data distribution and identified the variability level within each interval (between maximum and minimum observed values). Histograms representing the frequency distribution of available modalities among the studied material were plotted. Analysis of variance (ANOVA) was performed on genotypes as treatment with different replications. Principal component analysis (PCA) was performed to evaluate relationships among morphological traits and apricot accessions based on comparable properties. In addition, Pearson correlation coefficients and correspondence analysis were applied to identify putative redundancy and to discriminate relevant informative traits.
Results and discussion
Morphological variability assessment: The descriptive analysis revealed high variability for all studied characters and estimated by the interval of variation (median, maximum and minimum values) and the coefficient of variation (CV) (Table 3). The results revealed an important genotype impact on quantitative and qualitative variables. The quantitative descriptive analysis revealed a coefficients of variation (CV) between studied cultivars ranging from 11.39 to 37.27 % for LBL and FW respectively. However, the highest CV values were obtained for the qualitative characters, especially for petiole thickness (LPT: 62.9 %) and relative area of over colour of fruit (FROC: 61.39 %), while the stone size compared to fruit (SSCF) and the tree yield (TY) were in the order of 21.09 and 27.84 % respectively. These characters are summarized in Table 3. The analysis of variance (Table 3) revealed significant differences for all the examined quantitative traits,revealed a high level of variability in different morphological traits among all apricot accessions. All the characters were highly polymorphic for all studied Moroccan apricots. This result agreed with recent studies [13], such as the use of UPOV descriptors revealed high variability among apricot cultivars.
Modalities frequency of morphological traits: Frequency histograms plotted as a function of the different modalities of studied variables and number of accessions belonging to each modality are illustrated in Figure 2. Least frequency values were recorded for quantitative characters compared to qualitative ones (Figure 2). The leaf variables were highly represented and discriminated the differentiation between studied accessions. Leaf blade dimensions (LBL, LBW) had the same frequencies ranging from 0.05 to 0.2 with maximum values around 6 to 7 cm for most apricot accessions, whereas the frequency of the LBL / LBW ratio had a value of 1 which would explain the triangular shape of the leaves. Concerning fruit characters, the dimension’s variables (FLW, and FH/FLW) have a good quality of representation. This is explained by the small deviations found between median and mean values (3.66-3.69 cm), (3.11- 4.03 cm) and (0.92-1.12 cm) for FLW, FH and FH/FLW respectively, with frequency values around 0.2 to 0.3. However, the apricot cultivars were heterogeneous regarding fruit and stone weights with high frequencies (> 0.3) around the mean values of both variables, 34.84 g and 1.73 g respectively.
Regarding qualitative parameters (Figure 2), the most frequent values for the fruit shape traits (FSLV, FSVV and FFS) showed a predominance of a rounded fruit shape, the rectangular form shape was specific to other cultivars (30 %). The depth of suture (FDS) was slightly sunken associated with a medium pedicel cavity (FDPC). Fruit surface (FFS) was predominantly smooth and most fruits were mostly (98 %) asymmetric (FS). Studied apricots were characterized with average firmness of flesh (FFF): almost 90 % of fruits were not firm nor soft at commercial maturity. In addition, flesh and skin colours showed a significant variability of the analysed apricots, this is explained by the continuous distribution of the light to orange colour and sometimes red for some cultivars. The orange colour of flesh and of skin was the most common colour found in the 92 studied accessions. However, some other cultivars had intense orange and red colours.
Apricot accessions were shown to have higher frequencies and more homogeneity when qualitative parameters were used, compared to quantitative ones. Comparing these results with previous studies, the main differences between Moroccan apricots and European, Tunisian and Mexican ones were related to the fruit weight, color and dimensions. Moroccan apricots have medium-sized fruits, smaller than European apricots [13], larger than Mexican apricots [14] and slightly similar to Tunisian apricots [15]. Concerning the apricot shape, high variability was observed just as the study of [16] who reported that the rounded shape was the most dominant form. The market value of fruit size and shape are important physical attributes in sorting, sizing, packaging and transportation of fruits, and designing relevant equipment [17] reported that a high variability in some Serbian apricot selections evaluated based on properties related to fruit size and shape, and significant differences among selections were observed for all physical attributes. Moreover, significant variability was also reported for leaf dimensions and shapes since it covered almost all of the available modalities. This result was in agreement with previous ones reported by [18] who showed the importance of the apricot leaf characteristics in the identification and discrimination of genotypes.
The orange colour was the prevalent of the studied cultivars, which confirmed the results of previous studies [19] who reported a dominance of the orange coloured apricots compared to white, yellow and red, also this indicates that Moroccan apricots are rich in carotenoids [20] reported that carotenoid content in apricot fruit showed a good correlation with the skin and flesh colour, with apricots having orange coloured flesh containing higher levels of carotenoids than those with white-coloured flesh. This result was consistent with our previous study [21], we found that the development of an orange colour during apricot ripening was accompanied with the accumulation of carotenoids pigment. Indeed, the orange cultivars Marouch 16 (23.21 mg/g) was found to be the richest in total carotenoids, it was also rich in β-carotene (1491.79 µg/100 g) and Provitamin A (273.47 mg/100 g).
Correlations of morphological traits:Significant correlations were found between different characters related to the 34 polymorphic variables, they were estimated using the Pearson coefficient (Table 4). With regard to the quantitative variables, significant coefficient values were found, especially for variables related to fruit and stone weight and fruit dimensions (FW, SW, FH, FLW). The correlations found between fruit weight (FW), fruit lateral width (FLW) and height (FH) were high (0.909, 0.940 respectively), whereas the stone weight (SW) was positively correlated with FW (0.744), FH (0.748) and FLW (0.518) [22] found similar correlations, especially for variables related to fruit weight and dimensions. The correlations found between fruit weight (FW), fruit lateral width and height (FLW, FH) were high (0.929, 0.967 respectively) and reached 0.623 between fruit weight and stone weight (FW, FSW). The same correlation between fruit weight and dimensions was reported previously by [23]. Furthermore, fruit and stone weight (0.744) were found to be correlated with flesh firmness (0.507, 0.604 respectively), flesh colour (0.557; 0.816 respectively), un-blush skin color (0.690, 0.549 respectively) and blush skin color (0.825; 0.754 respectively).
Previous studies [24] also reported that fruit weight was significantly correlated with stone weight (r > 0.7), specifically larger fruits generally had larger stones. In fact, apricot stones have been used in genotype identification and have a high utilitarian value in characterization of genetic diversity [25] justified that apricot fruit weight is directly related to fruit length, width, fruit flesh and stone weight. Furthermore, the correlation between fruit length and stone weight was 0.61, since larger fruits harbour larger stones.
Leaf dimensions (LBL and LBW) were also significantly correlated (0.705), positively, with stone size (SSCF) and negatively with blade tip shape (LBST). The same correlation between leaf dimensions was reported previously by [26] who observed high correlation between the leaf blade length and width (LBL, LBW) (0.86), showing that the leaf blade increased relative to all dimensions during development. The intensity of over color (FIOC) and relative area of over color (FROC) variables were negatively correlated (-0.660). FIOC was positively correlated with flesh color (0.603), un-blush skin color (0.603), blush skin color (0.850), fruit surface (0.595), stone weight (0.566) and leaf blade undulation (0.547); and negatively correlated with symmetry in ventral view (-0.595) and number of glands (-0.557). On the other hand, FROC was positively correlated with the number of glands in the petiole (0.647) and negatively correlated with blush skin color (-0.622). Additionally, Tree yield (TY) was positively correlated with stone size (SSCF), leaf blade length (LBL)and petiole length (LPL); and negatively correlated with firmness of flesh (FFF) which was positively correlated with fruit weight (FW), stone weight (SW), fruit height (FH), shape in ventral view of fruit (FSVV), un-blush skin color (FUBC) and blush skin color (FBC).
The studied apricots were considered commercially ripe, they had reached full size, full color development and a very firm texture to withstand handling, transport and storage conditions. This explains the positive correlations found between the fruit weight, stone weight, fruit dimensions, flesh firmness and colors. It is worth noting the existence of independent variables with a null Pearson correlation coefficient (Table 3) such as correlations of firmness of flesh (FFF) with apex shape (FAS), as well as symmetry in ventral view (FS) and fruit surface (FFS) variables. Regarding fruit and leaf correlations, the apricot leaf traits can better discriminate between the 92 accessions compared to fruit ones.
Traits involved in morphological differentiation: Principal component analysis (PCA) was performed to identify the principal distinguishing characters of the variability. More than 48 % of the total variance among accessions was explained by the first two components (PC1 and PC2). PC1 discriminates mainly fruit characters (Fig. 2a), such as fruit and stone weight (FW, SW), fruit dimensions (FH, FLW) and fruit colours (FUBC, FBC, FFC, FIOC, FROC) and accounts for 32.21 % of the total variance. While PC2 explained 15.89 % of variability and was represented especially by leaf dimensions (LBL, LBW, LPL), leaf blade form (LBST, LBIM) and fruit apex shape (FAS).
Positive values of PC1 and PC2 were specific to the quantitative characters while negative part, like to axis origin, were specific to the qualitative variables. The plotted data indicated the presence of an important variability between the apricot accessions (Figure 2a). Regarding dispersion of apricot accessions on diagram (Figure 2b) and variables / PCA component correlations, it was worth noting that morphometric variables, especially of leaf traits, were mostly involved in the observed germplasm variability. In addition, it should be noted that according to the correlations between variables and both main components, we can observe a heterogeneity between studied accessions linked mainly to the qualitative compared to the quantitative characters.
When considering the geographical origin of the accessions, the results show that the geographic region has not an impact on apricot diversity. Indeed, a structural analysis seems important to illustrate and to demonstrate this geographical effect.
Figure 2: Representation the 92 Moroccan apricot accessions according to their morphological characters.
(a) Representation of variables according to their PCA contributions on the two main axes.
(b) Segregation of the 92 apricot accessions according their variables.
(Explanations of the word abbreviations are shown in Table 1)
Figure 3: Hierarchical clustering of the 92 apricot accessions based on Ward’s method using Euclidian distance with standard deviation.
Variability structure of Moroccan apricots:All PCA components were plotted in a dendrogram structure (Figure 3). The obtained results from hierarchical clustering, using the linkage method between groups, distinguished all the 92 accessions and revealed two separate clusters: I and II (Figure 3). The first group (I) encompassed 89 cultivars, while the Boum A1, Boum A2 and SKT 5 constituted the second cluster (II).
Cluster I included apricots with medium to large fruit and cluster II included apricots with large leaf size. At a dissimilarity level of 21, cluster I could be divided into two subgroups: IA; 81 cultivars which in turn subdivide to other heterogeneous subgroups and IB; 8 cultivars (Outat 1, Outat 2, RK1, RK2, RK3, RT1, Outat 6, Clone C, RT4). The 81 cultuvars are characterized by colored (orange and red) apricots and large fruits size, they included genotypes of INRAM group mainly Gelitano and Marouch group (3, 4 and 16). In addition to INRAM accessions, this population was also composed with apricots issued from Moulouya valley (OutatElhaj). These apricots are grown under arid conditions with a relatively long winter period and lower temperatures [23-26]. These accessions display higher chilling requirements than the first group and are characterized by oval-shaped, relatively large-sized fruit with yellow flesh, similar to the fruits of Canino [27]. The apricots of second subgroup (IB: 8 cultivars) have a common point regarding a small size of fruits, a light colour and the majority of these accessions were issued from Ziz valley. They are grown under Saharan climatic conditions with a short winter period and high temperatures starting in early spring, and they are characterized by having low chilling requirements and are early blooming, with flat-shaped, white flesh fruit; they, however, display a limited ability for fresh fruit conservation and processing.
This classification structure was able to show that the dimensions of the leaf and the fruit of apricot as well as the colour of the fruit represent the discriminating parameters of the genetic diversity of the studied apricot germplasm. The morphometry of more than 90 % of the studied cultivars was based on a balanced equation between dimensional and colour variables. The other apricot cultivars represent less than 10 % of the totality, these were mainly cultivars with large leaves size, especially Boum LG3, Agdez LG1, Ab 3, Km 8 and SKT 5.
The obtained classification (Figure 3) was not in accordance with the geographic origin of apricot accessions compared to genotype factor. No clear morphological structure was displayed according to the geographical origin of accessions nor with their environment source. This is in agreement with the previous study by [28], who analyzed different apricot cultivars and concluded that varieties belonging to the same geographic population could not be grouped into a same group. It is also worth noting that the apricot genotype had a marked effect on the observed variability and its structure, particularly on morphological characters of fruits and leaves that highlight an important genetic diversity useful for the development of the apricot sector both in Morocco and in the Mediterranean region. The large variation in apricot fruits is a particularly important performance for breeding of larges cultivars for better marketability and distribution to the growers.
In addition, regarding our results, the assessed Moroccan apricot germplasm presented great variability, related to genotype, comparable to that of European [29-35]. Indeed, different results obtained by different analyses undergone by this study support the hypothesis that Moroccan apricots have a genetic lineage distinct from other available germplasm. Originally, the Moroccan apricots were multiplied by seeds, with the exception of some cultivars [36-41]. As a consequence, the multiplication of seeds, unlike the propagation of the graft dose not fixe the genotype, leads to many successive breeding events that generate genetic drift within a population. This evolutionary model involves both high genetic differentiation in apricot populations and low genetic diversity. This has been confirmed by previous genetic studies on Moroccan and Tunisian apricots [41-45].
Acknowledgments
This work was supported by a grant from the Moroccan ministry of higher education, scientific research including professional training for Jamal Ayour. The authors would like to thank the National Institute of Agricultural Research (INRA), especially the Agronomic Regional Center in Marrakesh (Morocco). The authors also thank Dr HasnaâHarrak, from the Laboratory of Food Technology and Quality, Research Unit of Plants improvement and Quality (INRA), for her precious collaboration and contribution.
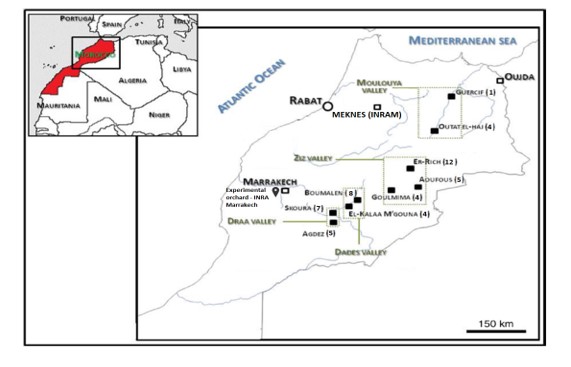
Figure 1: Geographic origin of studied accessions in Morocco.
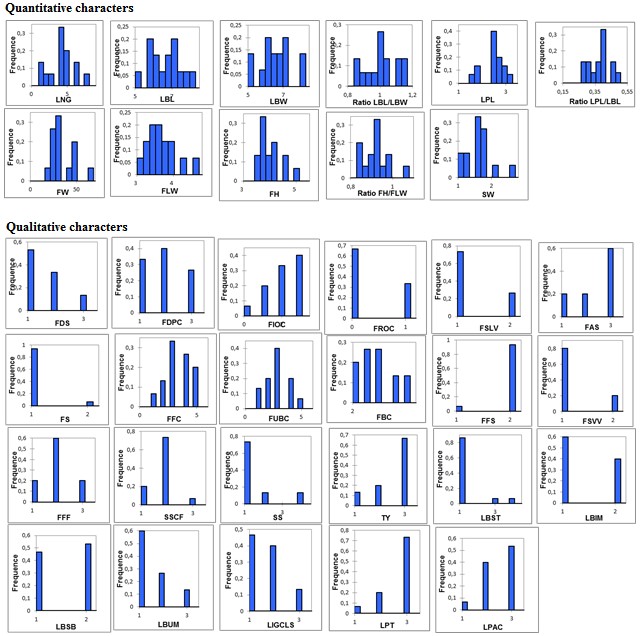
Figure 2:Frequency histograms based on the morphological of 11 quantitative and 23 qualitative apricot characters. (Explanations of the word abbreviations are shown in Table 1).
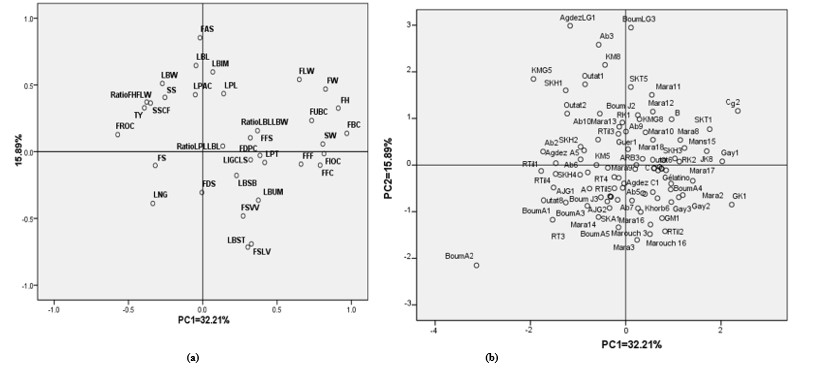
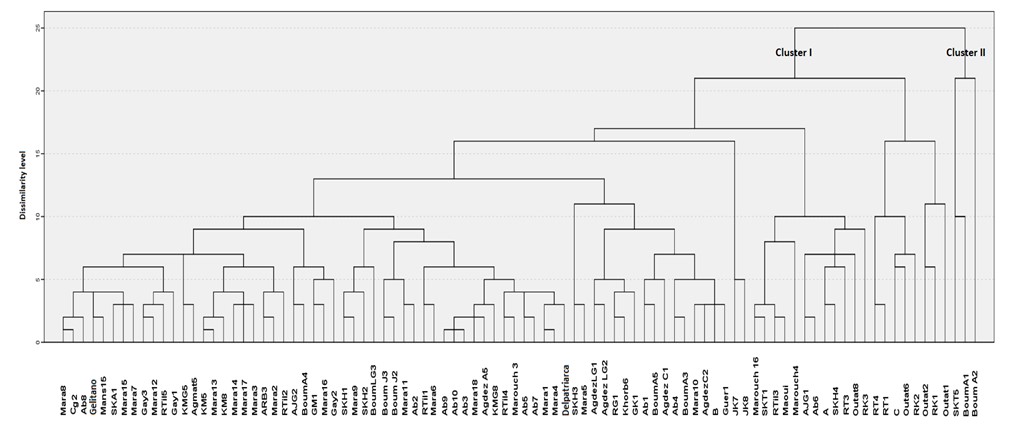
Figure 3: Hierarchical clustering of the 92 apricot accessions based on Ward’s method using Euclidian distance with standard deviation.
|
Name |
Geographical site |
Geographical region |
|
Agdez A5 |
Agdez |
Drâa valley |
|
Agdez C1 |
Agdez |
Drâa valley |
|
Agdez LG2 |
Agdez |
Drâa valley |
|
AgdezC2 |
Agdez |
Drâa valley |
|
AgdezLG1 |
Agdez |
Drâa valley |
|
SKA1 |
Skoura |
Drâa valley |
|
SKH1 |
Skoura |
Drâa valley |
|
SKH2 |
Skoura |
Drâa valley |
|
SKH3 |
Skoura |
Drâa valley |
|
SKH4 |
Skoura |
Drâa valley |
|
SKT1 |
Skoura |
Drâa valley |
|
SKT5 |
Skoura |
Drâa valley |
|
KM5 |
KalaatMeggouna |
Dadès valley |
|
KM8 |
KalaatMeggouna |
Dadès valley |
|
KMG5 |
KalaatMeggouna |
Dadès valley |
|
KMG8 |
KalaatMeggouna |
Dadès valley |
|
Boum A2 |
Boumalen |
Dadès valley |
|
Boum J2 |
Boumalen |
Dadès valley |
|
Boum J3 |
Boumalen |
Dadès valley |
|
BoumA1 |
Boumalen |
Dadès valley |
|
BoumA3 |
Boumalen |
Dadès valley |
|
BoumA4 |
Boumalen |
Dadès valley |
|
BoumA5 |
Boumalen |
Dadès valley |
|
BoumLG3 |
Boumalen |
Dadès valley |
|
Outat1 |
OutatElhaj |
Moulouya valley |
|
Outat2 |
OutatElhaj |
Moulouya valley |
|
Outat6 |
OutatElhaj |
Moulouya valley |
|
Outat8 |
OutatElhaj |
Moulouya valley |
|
Guer1 |
Guersif |
Moulouya valley |
|
AJG1 |
Aoufous |
Ziz valley |
|
AJG2 |
Aoufous |
Ziz valley |
|
AJRB3 |
Aoufous |
Ziz valley |
|
JK7 |
Aoufous |
Ziz valley |
|
JK8 |
Aoufous |
Ziz valley |
|
Gay1 |
Goulmima |
Ziz valley |
|
Gay2 |
Goulmima |
Ziz valley |
|
Gay3 |
Goulmima |
Ziz valley |
|
GM1 |
Goulmima |
Ziz valley |
|
RG1 |
Er-Rich |
Ziz valley |
|
RK1 |
Er-Rich |
Ziz valley |
|
RK2 |
Er-Rich |
Ziz valley |
|
RK3 |
Er-Rich |
Ziz valley |
|
RT1 |
Er-Rich |
Ziz valley |
|
RT3 |
Er-Rich |
Ziz valley |
|
RT4 |
Er-Rich |
Ziz valley |
|
RTil1 |
Er-Rich |
Ziz valley |
|
RTil2 |
Er-Rich |
Ziz valley |
|
RTil3 |
Er-Rich |
Ziz valley |
|
RTil4 |
Er-Rich |
Ziz valley |
|
RTil5 |
Er-Rich |
Ziz valley |
|
Name |
Geographical site |
Geographical region |
|
Agmat5 |
INRAM |
INRAM |
|
Khorb6 |
INRAM |
INRAM |
|
Mans15 |
INRAM |
INRAM |
|
Maoui |
INRAM |
INRAM |
|
Delpatriarca |
INRAM |
INRAM |
|
Gelitano |
INRAM |
INRAM |
|
Marouch3 |
INRAM |
INRAM |
|
Marouch4 |
INRAM |
INRAM |
|
Marouch16 |
INRAM |
INRAM |
|
GK1 |
Marrakech |
Marrakech |
|
Cg2 |
Marrakech |
Marrakech |
|
A |
Marrakech |
Marrakech |
|
B |
Marrakech |
Marrakech |
|
C |
Marrakech |
Marrakech |
|
Ab1 |
Marrakech |
Marrakech |
|
Ab2 |
Marrakech |
Marrakech |
|
Ab3 |
Marrakech |
Marrakech |
|
Ab4 |
Marrakech |
Marrakech |
|
Ab5 |
Marrakech |
Marrakech |
|
Ab6 |
Marrakech |
Marrakech |
|
Ab7 |
Marrakech |
Marrakech |
|
Ab8 |
Marrakech |
Marrakech |
|
Ab9 |
Marrakech |
Marrakech |
|
Ab10 |
Marrakech |
Marrakech |
|
Mara1 |
Marrakech |
Marrakech |
|
Mara2 |
Marrakech |
Marrakech |
|
Mara3 |
Marrakech |
Marrakech |
|
Mara4 |
Marrakech |
Marrakech |
|
Mara5 |
Marrakech |
Marrakech |
|
Mara6 |
Marrakech |
Marrakech |
|
Mara7 |
Marrakech |
Marrakech |
|
Mara8 |
Marrakech |
Marrakech |
|
Mara9 |
Marrakech |
Marrakech |
|
Mara10 |
Marrakech |
Marrakech |
|
Mara11 |
Marrakech |
Marrakech |
|
Mara12 |
Marrakech |
Marrakech |
|
Mara13 |
Marrakech |
Marrakech |
|
Mara14 |
Marrakech |
Marrakech |
|
Mara15 |
Marrakech |
Marrakech |
|
Mara16 |
Marrakech |
Marrakech |
|
Mara17 |
Marrakech |
Marrakech |
|
Mara18 |
Marrakech |
Marrakech |
|
Organs |
Characters and modalities |
|
|
Leaf |
Leaf blade |
Length (LBL) (cm) |
|
Width (LBW) (cm) |
||
|
Petiole |
Length (LPL) (cm) |
|
|
Number of glands (LNG) |
||
|
Ratios
|
Leaf blade length/Leaf blade width (LBL/LBW) |
|
|
Petiole length/blade length (LPL/LBL) |
||
|
Leaf blade: undulation of margin (LBUM) |
1: slight, 2: medium, 3: strong |
|
|
Intensity of green color of upper side (LIGCLS) |
1: light, 2: medium, 3: dark |
|
|
Leaf blade: shape of base (LBSB) |
1: attenuate, 2: truncate, 3: subcordate, 4: cordate |
|
|
Leaf blade: shape of tip (LBST) |
1: mucronate, 2: cuspidate, 3: acuminate |
|
|
Leaf blade: angle of tip (LBAT) |
1: narrow acute, 2: broad acute, 3: obtuse |
|
|
Leaf blade: incisions on margin (LBIM) |
1: crenate, 2: bicrenate, 3: serrate, 4: biserrate |
|
|
Petiole thickness (LPT) |
1: small, 2: medium, 3: large |
|
|
Petiole anthocyanin coloration (LPAC) |
1: weak, 2: medium, 3: strong |
|
|
Fruit |
Weight |
Fruit weight (FW) (g) |
|
Dimensions |
Lateral width (FLW) (cm) |
|
|
Ventral width (FVW) (cm) |
||
|
Height (FH) (cm) |
||
|
Ratios
|
Fruit lateral width/Fruit ventral width (FLW/FVW) |
|
|
Fruit height/Fruit ventral width (FH/FVW) |
||
|
Depth of suture (FDS) |
1: slightly sunken, 2: moderately sunken, 3: deeply sunken |
|
|
Depth of pedicel cavity (FDPC) |
1: shallow, 2: medium, 3: deep |
|
|
Intensity of over color (FIOC) |
1: weak, 2: medium, 3: strong, 4: absent |
|
|
Relative area of over color (FROC) |
1: small, 2: medium, 3: large, 4: absent |
|
|
Firmness of flesh (FFF) |
1: soft, 2: medium, 3: firm |
|
|
Shape in lateral (or profile) view (FSLV) |
1: rounded, 2: triangular, 3: trapezoidal, 4: rectangular |
|
|
Shape in ventral (or frontal) view (FSVV) |
1: rounded, 2: triangular, 3: trapezoidal, 4: rectangular |
|
|
Apex shape (FAS) |
1: depressed, 2: flat, 3: rounded, 4: pointed |
|
|
Symmetry in ventral view (FS) |
1: asymmetric, 2: symmetric |
|
|
Fruit surface (FFS) |
1: smooth, 2: bumpy shoots, 3: predominantly on one-year-old shoots |
|
|
Flesh color (FFC) |
1: light, 2: cream to yellow, 3: light orange, 4: medium orange, 5: dark orange |
|
|
Un-blush skin color (FUBC) |
1: light, 2: cream to yellow, 3: light orange, 4: orange, 5: dark orange |
|
|
Blush skin color (FBC) |
1: light, 2: cream to yellow, 3: light orange, 4: orange, 5: dark orange 6: red |
|
|
Stone |
Weight |
Stone weight (SW) (g) |
|
Size compared to fruit (SSCF) |
1: small, 2: medium, 3: large |
|
|
Shape (SS) |
1: oval, 2: round, 3: oblong, 4: elliptical |
|
|
Tree |
Yield: mean fruit yield per apricot tree (TY) |
1: few, 2: medium, 3: many |
|
- |
Variables |
Mean |
SD |
Median |
Maximum |
Minimum |
CV% |
Fisher |
P > F |
|
Quantitative variables |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
Leaf |
Leaf blade length (LBL) (cm) |
6.8 |
0.77 |
6.8 |
8.5 |
5 |
11.39 |
19 |
<0.001 |
|
Leaf blade width (LBW) (cm) |
6.59 |
0.92 |
6.7 |
8.5 |
3.7 |
13.94 |
11 |
<0.001 |
|
|
Ratio LBL/LBW |
1.04 |
0.11 |
1.01 |
1.57 |
0.82 |
10.96 |
23 |
<0.001 |
|
|
Petiole length(LPL) (cm) |
2.64 |
0.48 |
2.6 |
4.3 |
1.3 |
18.06 |
44 |
<0.001 |
|
|
Ratio LPL/LBL |
0.39 |
0.07 |
0.39 |
0.6 |
0.22 |
17.63 |
44 |
<0.001 |
|
|
Number of glands in petiole |
4.02 |
1.3 |
4 |
8 |
1 |
32.21 |
61 |
<0.001 |
|
|
Fruit |
Fruit weight (g) |
34.84 |
12.99 |
28.51 |
67.23 |
19.72 |
37.27 |
68 |
<0.001 |
|
Lateral width (cm) |
3,69 |
0.39 |
3.66 |
4.61 |
2.13 |
10.8 |
17 |
<0.001 |
|
|
Height (cm) |
4.03 |
0.42 |
3.11 |
5.02 |
1.59 |
10.38 |
17 |
<0.001 |
|
|
Ratio FH/FLW |
1.12 |
1.06 |
0.92 |
1.07 |
0.84 |
6.73 |
8 |
<0.001 |
|
|
Stone |
Stone weight (g) |
1.73 |
0.38 |
1.48 |
2.87 |
0.82 |
21.74 |
48 |
<0.001 |
|
Qualitative variables |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
Leaf |
Leaf blade: undulation of margin |
2 |
0.67 |
1 |
3 |
1 |
43.87 |
- |
- |
|
Leaf blade: incisions on margin |
1 |
0.46 |
1 |
2 |
1 |
35.51 |
- |
- |
|
|
Leaf blade: shape of base |
2 |
0.5 |
2 |
2 |
1 |
35.51 |
- |
- |
|
|
Leaf blade: shape of tip |
1 |
0.81 |
1 |
4 |
1 |
33.52 |
- |
- |
|
|
Intensity of green color of upper side |
2 |
0.81 |
2 |
3 |
1 |
43.23 |
- |
- |
|
|
Petiole anthocyanin coloration |
2 |
0.72 |
3 |
3 |
1 |
42.06 |
- |
- |
|
|
Petiolethickness |
3 |
0.52 |
3 |
3 |
1 |
62.9 |
- |
- |
|
|
Fruit |
Depth of suture |
2 |
0.74 |
1 |
3 |
1 |
46.05 |
- |
- |
|
Depth of pedicel cavity |
2 |
0.8 |
2 |
3 |
1 |
41.32 |
- |
- |
|
|
Intensity of over color |
2 |
0.96 |
2 |
3 |
0 |
46.51 |
- |
- |
|
|
Relative area of over color |
0 |
0.49 |
0 |
1 |
0 |
61.39 |
- |
- |
|
|
Firmness of flesh |
2 |
0.14 |
2 |
3 |
2 |
17.03 |
- |
- |
|
|
Shape in lateral (or profile) view |
1 |
0.46 |
1 |
2 |
1 |
36.14 |
- |
- |
|
|
Shape in ventral (or frontal) view |
1 |
0.41 |
1 |
2 |
1 |
34.5 |
- |
- |
|
|
Apex shape |
2 |
0.83 |
3 |
3 |
1 |
34.5 |
- |
- |
|
|
Symmetry in ventral view |
1 |
0.26 |
1 |
2 |
1 |
24.21 |
- |
- |
|
|
Fruit surface |
2 |
0.26 |
2 |
2 |
1 |
13.36 |
- |
- |
|
|
Fleshcolor |
3 |
1.18 |
3 |
5 |
1 |
34.8 |
- |
- |
|
|
Un-blush skin color |
3 |
1.23 |
3 |
5 |
1 |
36 |
- |
- |
|
|
Blush skin color |
4 |
1.51 |
3 |
6 |
1 |
42.61 |
- |
- |
|
|
Stone |
Size compared to fruit |
2 |
0.52 |
2 |
3 |
1 |
21.09 |
- |
- |
|
Shape |
2 |
0.34 |
2 |
4 |
1 |
14.88 |
- |
- |
|
|
Tree |
Yield |
2 |
0.69 |
2 |
3 |
1 |
27.84 |
- |
- |
Table 3:Descriptive statistics related to the 34 polymorphic morphological variables among the 92 accessions and analysis of variance (ANOVA) for quantitative variables (SD: standard deviation, CV: coefficient of variation, Fisher test values and probability level at a threshold of 0.05%)
|
|
FW |
FLW |
FH |
Ratio FH/FLW |
FDS |
FDPC |
FIOC |
FROC |
FSLV |
FSVV |
FAS |
FS |
FFC |
FUBC |
|
FW |
1 |
0.909 |
0.94 |
0.008 |
-0.053 |
0.309 |
0.581 |
-0.327 |
-0.073 |
0.003 |
0.434 |
-0.322 |
0.557 |
0.69 |
|
FLW |
- |
1 |
0.815 |
0.362 |
0.176 |
0.319 |
0.389 |
-0.184 |
-0.217 |
-0.106 |
0.53 |
-0.254 |
0.315 |
0.495 |
|
FH |
- |
- |
1 |
-0.243 |
-0.085 |
0.428 |
0.686 |
-0.358 |
0.106 |
0.121 |
0.271 |
-0.281 |
0.684 |
0.756 |
|
Ratio FH/FLW |
- |
- |
- |
1 |
0.443 |
-0.17 |
-0.452 |
0.277 |
-0.504 |
-0.344 |
0.459 |
0.023 |
-0.576 |
-0.39 |
|
FDS |
- |
- |
- |
- |
1 |
0.437 |
-0.061 |
0.199 |
0.127 |
0.047 |
-0.07 |
0.15 |
-0.049 |
-0.327 |
|
FDPC |
- |
- |
- |
- |
- |
1 |
0.099 |
0.244 |
0.247 |
0.259 |
0.043 |
0.369 |
0.484 |
0.148 |
|
FIOC |
- |
- |
- |
- |
- |
- |
1 |
-0.66 |
0.281 |
0.144 |
-0.215 |
-0.595 |
0.603 |
0.603 |
|
FROC |
- |
- |
- |
- |
- |
- |
- |
1 |
-0.107 |
0 |
0.177 |
0.378 |
-0.495 |
-0.434 |
|
FSLV |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
0.829 |
-0.678 |
-0.161 |
0.317 |
0.074 |
|
FSVV |
- |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
-0.458 |
-0.134 |
0.117 |
0.215 |
|
FAS |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
0.2 |
-0.029 |
0.138 |
|
FS |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
0.14 |
-0.459 |
|
FFC |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
0.472 |
|
FUBC |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
|
FBC |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
FFS |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
FFF |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
SSCF |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
SW |
- |
- |
- |
- |
- |
- |
|
- |
- |
- |
- |
- |
- |
- |
|
SS |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
TY |
|
- |
- |
- |
- |
- |
- |
- |
-- |
- |
- |
- |
|
- |
|
LBST |
- |
- |
- |
|
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
LBIM |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
LBSB |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
LBUM |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
LIGCLS |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
LPT |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
LPAC |
- |
- |
- |
- |
- |
- |
|
- |
- |
- |
- |
- |
- |
- |
|
LNG |
- |
- |
- |
- |
- |
- |
|
- |
- |
- |
- |
- |
- |
- |
|
LBL |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
LBW |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
Ratio LBL/LBW |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
LPL |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
Ratio LPL/LBL |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
- |
LBIM |
LBSB |
LBUM |
LIGCLS |
LPT |
LPAC |
LNG |
LBL |
LBW |
Ratio LBL/LBW |
LPL |
Ratio LPL/LBL |
- |
- |
|
FW |
0.306 |
0.005 |
0.077 |
0.219 |
0.383 |
0.113 |
-0.363 |
0.119 |
-0.116 |
0.367 |
0.307 |
0.213 |
- |
- |
|
FLW |
0.423 |
-0.152 |
0.029 |
0.24 |
0.203 |
0.123 |
-0.408 |
0.12 |
-0.153 |
0.389 |
0.344 |
0.279 |
- |
- |
|
FH |
0.352 |
0.207 |
0.16 |
0.302 |
0.446 |
0.082 |
-0.303 |
0.155 |
-0.096 |
0.396 |
0.257 |
0.139 |
- |
- |
|
Ratio FH/FLW |
0.121 |
-0.583 |
-0.177 |
-0.038 |
-0.387 |
0.091 |
-0.18 |
-0.055 |
-0.084 |
-0.012 |
0.192 |
0.278 |
- |
- |
|
FDS |
-0.115 |
0.225 |
0.157 |
0.268 |
0.157 |
-0.03 |
0.144 |
-0.336 |
-0.434 |
0.111 |
0.013 |
0.239 |
- |
- |
|
FDPC |
0.423 |
0.612 |
-0.176 |
0.329 |
0.676 |
-0.075 |
0.244 |
0.177 |
-0.121 |
0.403 |
0.172 |
0.02 |
- |
- |
|
FIOC |
-0.205 |
0.211 |
0.547 |
0.034 |
0.161 |
0.062 |
-0.557 |
0.132 |
-0.121 |
0.372 |
0.065 |
-0.049 |
- |
- |
|
FROC |
0.289 |
0.094 |
-0.328 |
0.135 |
0.158 |
0.152 |
0.647 |
0.23 |
0.16 |
0.071 |
0.2 |
0.074 |
- |
- |
|
FSLV |
-0.492 |
0.262 |
0.392 |
0.287 |
0.337 |
-0.211 |
0.204 |
-0.228 |
-0.106 |
-0.098 |
0.076 |
0.214 |
- |
- |
|
FSVV |
-0.408 |
0.134 |
0.093 |
0.477 |
0.28 |
-0.108 |
0.238 |
-0.076 |
0.175 |
-0.285 |
-0.067 |
-0.074 |
- |
- |
|
FAS |
0.612 |
-0.033 |
-0.371 |
0.238 |
0 |
0.431 |
-0.128 |
0.33 |
0.376 |
-0.057 |
0.417 |
0.222 |
- |
- |
|
FS |
0.327 |
0.25 |
-0.199 |
0.127 |
0.149 |
-0.202 |
0.568 |
-0.019 |
-0.039 |
0.017 |
0.064 |
0.068 |
- |
- |
|
FFC |
0.071 |
0.444 |
0.309 |
0.167 |
0.489 |
-0.264 |
-0.058 |
0.012 |
-0.22 |
0.378 |
0.201 |
0.165 |
- |
- |
|
FUBC |
0.225 |
0.131 |
-0.08 |
0.38 |
0.24 |
-0.007 |
-0.193 |
0.122 |
0.116 |
0.057 |
-0.18 |
-0.297 |
- |
- |
|
FBC |
0.063 |
0.221 |
0.37 |
0.345 |
0.318 |
0.072 |
-0.468 |
0.091 |
-0.095 |
0.307 |
0.216 |
0.121 |
- |
- |
|
FFS |
-0.327 |
-0.25 |
0.199 |
-0.127 |
-0.149 |
0.202 |
-0.568 |
0.019 |
0.039 |
-0.017 |
-0.064 |
-0.068 |
- |
- |
|
FFF |
0 |
0 |
0 |
0.452 |
0.177 |
-0.17 |
-0.116 |
-0.098 |
-0.052 |
-0.021 |
-0.165 |
-0.191 |
- |
- |
|
SSCF |
0.218 |
0.286 |
-0.36 |
-0.319 |
0.075 |
0.202 |
0.02 |
0.621 |
0.666 |
-0.081 |
0.175 |
-0.214 |
- |
- |
|
SW |
0.029 |
0.054 |
0.224 |
0.047 |
0.262 |
-0.34 |
-0.143 |
-0.066 |
-0.343 |
0.441 |
0.128 |
0.131 |
- |
- |
|
SS |
-0.027 |
-0.165 |
-0.296 |
-0.124 |
-0.036 |
-0.077 |
0.105 |
0.349 |
0.367 |
-0.04 |
0.037 |
-0.157 |
- |
- |
|
TY |
-0.227 |
-0.05 |
-0.034 |
-0.443 |
-0.208 |
0.19 |
-0.207 |
0.538 |
0.466 |
0.077 |
0.509 |
0.198 |
- |
- |
|
LBST |
-0.313 |
0.359 |
0.57 |
0.073 |
0.214 |
-0.289 |
0.056 |
-0.406 |
-0.538 |
0.205 |
-0.063 |
0.233 |
- |
- |
|
LBIM |
1 |
0.218 |
-0.417 |
0.195 |
0.228 |
0.264 |
0.015 |
0.321 |
0.126 |
0.241 |
0.194 |
0.017 |
- |
- |
|
LBSB |
- |
1 |
-0.05 |
0.319 |
0.598 |
0.274 |
0.289 |
0.024 |
0.061 |
-0.021 |
-0.014 |
-0.075 |
- |
- |
|
LBUM |
- |
- |
1 |
0.089 |
-0.052 |
0.04 |
-0.258 |
-0.138 |
-0.359 |
0.329 |
0.322 |
0.445 |
- |
- |
|
LIGCLS |
- |
- |
- |
1 |
0.213 |
0.206 |
0.297 |
-0.061 |
0.151 |
-0.228 |
0.121 |
0.147 |
- |
- |
|
LPT |
- |
- |
- |
- |
1 |
0.241 |
0.287 |
-0.099 |
-0.147 |
0.11 |
0.166 |
0.184 |
- |
- |
|
LPAC |
- |
- |
- |
- |
- |
1 |
-0.292 |
0.173 |
0.377 |
-0.281 |
0.324 |
0.207 |
- |
- |
|
LNG |
- |
- |
- |
- |
- |
- |
1 |
-0.227 |
-0.083 |
-0.16 |
-0.228 |
-0.095 |
- |
- |
|
LBL |
- |
- |
- |
- |
- |
- |
- |
1 |
0.705 |
0.359 |
0.431 |
-0.209 |
- |
- |
|
LBW |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
-0.404 |
0.208 |
-0.254 |
- |
- |
|
Ratio LBL/LBW |
- |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
0.331 |
0.118 |
- |
- |
|
LPL |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
0.785 |
- |
- |
|
Ratio LPL/LBL |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
- |
- |
Table 4: Pearson correlation matrix established among the 34 polymorphic morphological characters. (Explanations of the word abbreviations are shown in Table 1).
Citation:Ayour J, Alahyane A, Elateri I, Alfeddy MN, Benichou M (2019) Assessment of Environment and Genetic Impacts On the Variability of Morphological Traits of Apricot Germplasm from Various Genetic and Environmental Resources in Morocco. Open Acc J Agri Res: OAJAR-100025
